Hereditary Breast Cancer and the Identification of BRCA1 and BRCA2
There are several possible explanations for the observed tendency of breast cancer to cluster in families. Chance may cause a common disorder such as breast cancer to afflict more than one member of a kindred. Alternatively, shared environmental risk factor exposures or common sociocultural practices (e.g., late age at first childbirth) may be responsible.
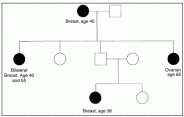
A common genetic background may increase familial risk by modulating sensitivity to environmental or reproductive risk factors in a similar fashion among members of a kindred. None of these hypotheses satisfactorily explain the 5-10% of breast cancer cases that occur in families that appear to be transmitting a predisposition to breast cancer in an autosomal dominant pattern (Fig. 11.1).
{kind=link}
In such families, breast cancer occurs in women of each generation, and approximately 50% of the female offspring of an affected woman are themselves affected. The disease tends to occur at an earlier age than in the population at large and is not infrequently bilateral.
There may be other types of cancer within the pedigree, especially ovarian cancer. Male breast cancer is not prominent in most families, but either males or females may transmit the predisposition. Early investigators suggested that the passage of a single autosomal dominant susceptibility allele could explain these uncommon families, if one accepted the assumption that expression of the susceptibility would be largely restricted to females. Unfortunately, until modern epidemiologic and molecular biologic techniques became available, this hypothesis was not proven.
In 1988, Newman et al provided the first formal demonstration (through segregation analysis) that one or more low-frequency, highly penetrant autosomal dominant alleles were responsible for a small proportion of breast cancers.
Subsequent studies indicated that one such locus was linked to the region of chromosome 17q21, and the term BRCA1 was coined to describe the putative gene. Clinical investigators observed that this locus was not only linked to hereditary breast cancer, but also to hereditary ovarian cancer.
This demonstration validated earlier clinical descriptions of an increased prevalence of ovarian cancer in families with an apparent predisposition to breast cancer. Families with an apparent predisposition to male breast cancer were not linked to the putative BRCA1 locus.
After an intensive multinational effort, BRCA1 was isolated by Miki et al in October of 1994. Shortly thereafter, germline mutations were detected in individuals from hereditary breast- and breast-ovarian cancer families.
The wild-type allele was shown to be lost in tumor tissue from patients with germline mutations, supporting the hypothesis that BRCA1 was a tumor-suppressor gene, analogous to P53 and RB, among others. In contradistinction to other tumor-suppressor genes, BRCA1 mutations could not be demonstrated in sporadic breast cancers. A number of hereditary breast and breast-ovarian cancer families could not be attributed to BRCA1 by either linkage or direct mutational analysis. Some of these families manifested an apparent predisposition to male breast cancer. By studying these families, a second susceptibility locus, called BRCA2, was mapped to chromosome 13q12.
In December 1995, the gene was cloned by Wooster et al and shown to be mutated in members of some, but not all, non-BRCA1 families.
Mark E. Robson
Breast conservation therapy for invasive breast cancer in Ashkenazi women with BRCA gene founder mutations. J Natl Cancer Inst 2003